Part 1
The Beginning of Life on Earth
Fossilized remains of microscopic life, date the beginning of life to approximately 3.8 billion years ago (though isotopic analysis of zircons could date life to as far back as 4.4 billion years ago.). Though there seems to be consensus on the when, the how life got its start remains a topic that has yet to have been agreed upon.
One of the first models to have been accepted (other than the belief not backed by any scientific evidence that life was created by god), is that life started from a primordial soup of the molecules necessary to form the building blocks of life (Herschy 2014). In this model molecules such as methane, ammonia, hydrogen gas, and water vapor were present in a solution and subjected to an electric shock that catalyzed their reaction to form amino acids, the building blocks of proteins.
A newer model, however, says life originated near hydrothermal vents deep in the ocean. These vents emit geothermally heated alkaline water, that when mixes with the oceans acidic seawater (the early oceans are thought to contain very high levels of CO2; making it acidic), provided the energy gradient necessary for the production of organic molecules, and then eventually, self-replicating molecules.
Despite these differing views on how life got its start, one thing that there seems to be broad consensus on is that life began as single cellular organisms in an environment antagonistic to the majority of life as we know it today. The earliest life forms were formed in an environment devoid of oxygen and high in methane. These life forms we know were microscopic and single cellular due to their fossilization in rocks that are dated to have been from 3.8 billion years ago. Then 2 billion years ago multicellular organism came into being. And from this point on, life will have evolved into its many complex forms we see today.
For many years, the process that drove the gradual change of life over billions of years remained a mystery. Then one man, Charles Darwin, discovered the mechanism that drives this change.
Natural Selection

Figure 1. Unknown author, Origin of Species. 1st edition, 1859, 1859, Public Domain
In 1859, Charles Darwin (1809-1882) published “On the Origin of Species” which detailed the newly discovered process called natural selection (Than 2018). In his book, he describes a process in which organisms more adapted to their surroundings are able to pass on their traits to their offspring and those not adopted to their surroundings are not. It is in this way that the great diversity observed between species originated over billions of years.
For natural selection to work, variation in the traits of a population of individuals that can be passed down to the organisms’ descendants must exist. These variations are then acted upon by the struggle of survival and it is this struggle that selects which traits are then able to be passed down to their descendants.
It is said that Darwin stumbled upon the theory of natural selection when in September of 1835, he visited the Galapagos Islands and observed a variety of small birds that, upon pondering in his return to London, realized that they were all different, but a closely related species.
Observations of two different mockingbird species of the Galapagos islands provided Darwin with the first clues of natural selection. However, the most famous and well-known animal of Darwin’s study remains the Finch.
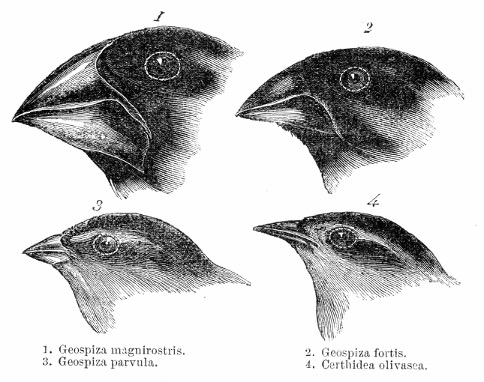
Darwin noticed that the Galapagos finches had variation in their beak sizes and shape and predicted that each finch descended from one original mainland species. He then called the process that selected for these differences in beaks natural selection. At the time, Darwin did not know about genetics. He was able to deduce the process, but not mechanism by which these processes succeeded. It was not until the 1900s that the gene will have been introduced as the unit of heredity and the full model of Natural selection will have been established (Mandal 2019).
The Gene: The Driving Force Behind Natural Selection
Charles Darwin established the fact that features of offspring are transferred from parent to offspring in 1859 in his publication of “On the Origin of Species”, but the mechanism by which these features were transferred where not yet known. Then with the research of Gregor Mendel, a 19th century Austrian monk and botanist, the mystery of the unit of heredity began to be unraveled.
Mendel studied and bred pea plants. In his studies he kept meticulous notes on the characteristics of each plants. He noticed that parental traits did not mix together in an offspring but appeared and disappeared through the generations with mathematical predictability. Scientist later then linked Mendel’s traits to a physical structure. This physical structure came to be known as the gene.
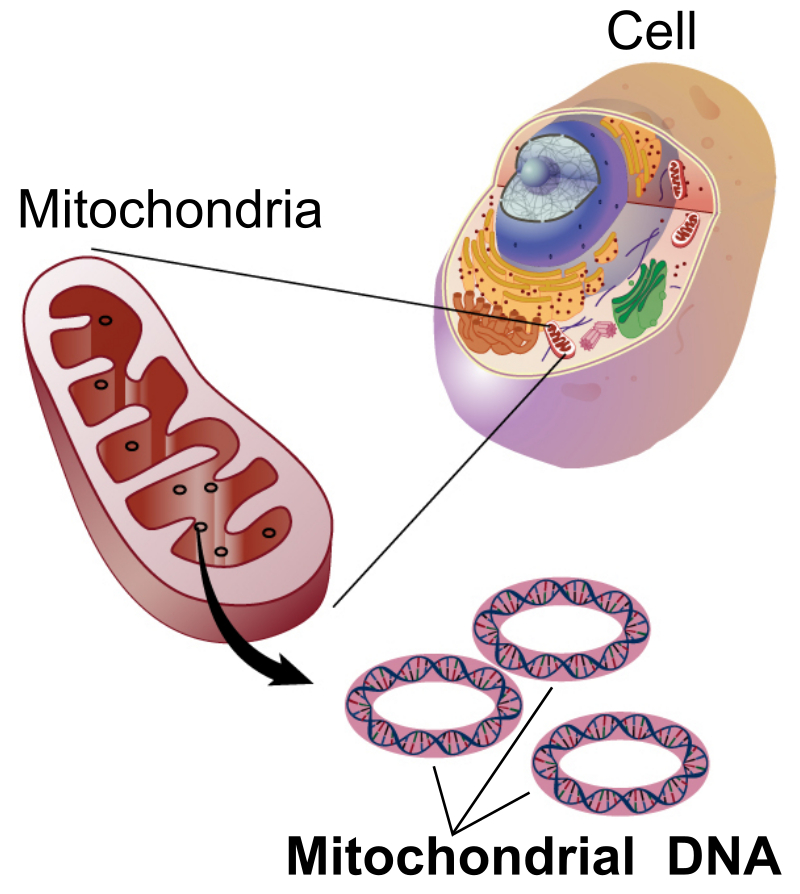
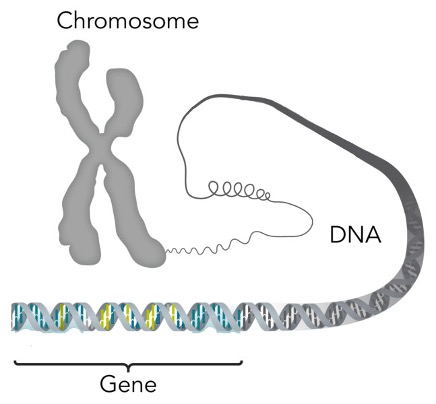
Defined as a basic physical and functional unit of heredity, genes are made up of deoxyribonucleic acid (DNA) and are located on chromosomes that reside in the nucleus of a cell, episomes, and mitochondria. Genes acts as a template to make molecules called proteins, the building blocks of life. It is through them, that organisms inherit their traits.
How do Organisms Inherit Genes?
Before speaking about the inheritance of genes, it is important to first discuss the cell. After all, it is through cells organisms inherit their genes. All living things are composed of cells. They provide structure to organisms, metabolizes food that is converted to energy, and carry out a myriad of other specialized functions. Cells have many parts each with a different function, but for the purposes of this paper, we will only focus on the nucleus. Typically, the nucleus of a cell will be located in proximity of its center. It is here that the chromosome is located.
Multicellular and single cellular organisms, when they reproduce, are essentially copying their cells and dividing in two to create a new organism. During cell division, the process in which genes are passed down from generation to generation, the contents of the cell will be broken down and all but the contents in the nucleus disappear. The extranuclear components are thought to be not transmitted from cell to cell. Nuclear contents, the chromosomes, are duplicated then segregated to two poles. The cell then cleaves itself in two and it is this way that chromosomes and, by extension, genes are passed on from organism to organism in a nuclear inheritance model.
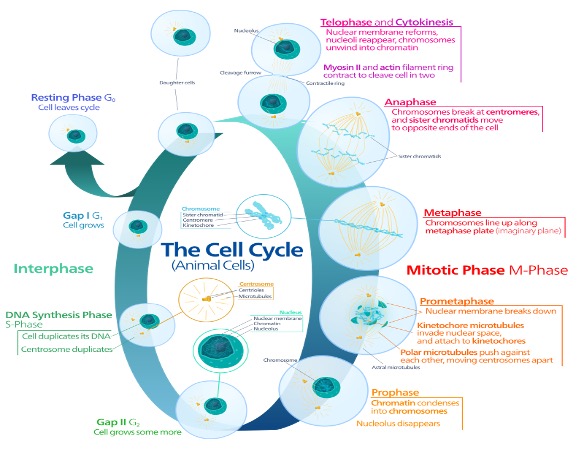
It is through this model that we typically tend to imagine the inheritance of genes. The gene, a stable unit located in each chromosome, is passed down from parent to offspring in nuclear inheritance model.
However, we now know of at least one other way that genes are passed on from offspring to offspring. And that is through cytoplasmic inheritance.
Part 2
Cytoplasmic Inheritance
It was important to first establish the background of the origin of life and the processes and structures that govern life to fully understand and think about cytoplasmic inheritance. Know that life has existed for almost four billion years. Know that all life, from human to bacteria, all arise from the process of natural selection through life that existed these four billion years ago. The variation that we see in the different organisms of a population, all arise from a process called natural selection, in which variation in a population is selected for and reproduced to cope with the stresses of existence. These variations are caused by genes. These genes are located in the nucleus of organisms and passed on from generation to generation through a nuclear inheritance. This is the model that has wide consensus among the scientific community. But what we find out is that this model is incomplete.
It is true that genes are held in the nucleus of cells and passed down from generation to generation, but what has also come to light is the fact that genes also exist outside of the nucleus and these genes are also sometimes transferred from parent to offspring through a model known as Cytoplasmic Inheritance. Cytoplasmic inheritance, or extranuclear inheritance, involves the transmission of genes that occur outside the nucleus. It is found in most eukaryotes and is commonly known to occur in cytoplasmic organelles such as mitochondria and chloroplasts or from cellular parasites such as viruses and bacteria.
The Discovery of Cytoplasmic Inheritance
In 1909, shortly after the wide acceptance of Gregory Mendel’s principles of inheritance. Carl Correns, a German botanist, and geneticist notice a strange pattern of inheritance in the plant Mirabilis jalapa (also known as the four o’clock flower).
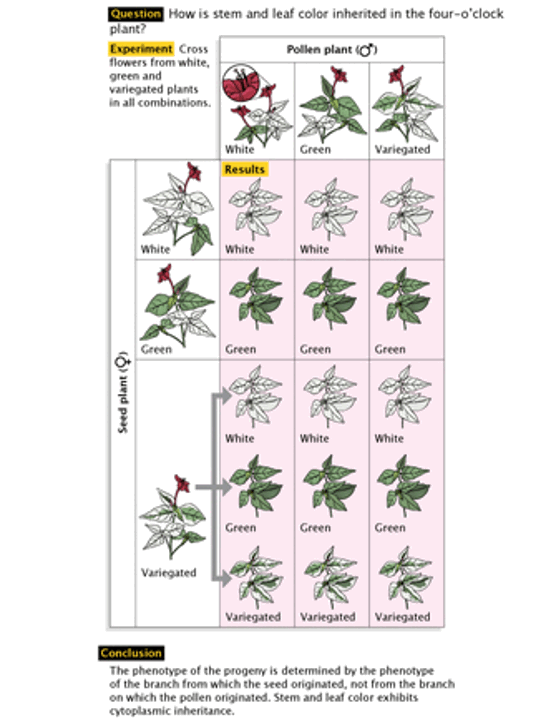
Correns notices that the leaves and stems on some plant would appear variegated, meaning that instead of being a single color, they would show multiple colors in a swirl like pattern, and within the same plant, branches may show a variety of leaf patterns (Miko 2008).. Seeds stemming from non-variegated plants would never produce variegated progeny, even when pollen came from a variegated flower. If seeds came from a variegated plant, the progeny would appear with either green, white, or variegated leaves, but never in a ratio that fit Mendel’s mathematical model. Furthermore, in doing cross-pollination experiments, Correns notices that leaf color in mirabilis were passes on uniparentally. Correns thus began to suspect that the maternal parent determined the leaf color phenotype because the paternal source of pollen never affected the outcome. In other words, Correns hypothesized that leaf color in Mirabilis was passed on uniparentally (Miko 2008)..
Around the same time, Erwin Baur separately stumbles upon an inheritance mode that deviated from the nuclear inheritance model. Baur noted in variegated Pelargonium zonale, green and white sections differed genetically in their plastids. One type of plastid expresses a green phonotype and the other a white phenotype. He then suggested that, in plant species with a biparental mode of inheritance for plasmids, small organelles that themselves contain DNA, RNA, or both. Zygotes of variegated plants developed with two distinct line of plastids. He theorizes that plasmids are randomly sorted in the course of cell division and this is the reason for areas of the plants that are pure green and areas that are pure white. And thus, the model of extranuclear inheritance began to be formed. But before we can conclusively say organisms exhibit cytoplasmic inheritance a few different phenomena and entities must be accounted for and excluded.
Establishing the Cytoplasmic Inheritance Model
During sexual reproduction of multicellular organisms, the female and gamete cell will fuse together, and this is the basis of formation of a new organism. Generally, this new organism will inherit one set of chromosomes from each parent and located on these chromosomes are genes. It was previously thought that this was the extent of which genes are inherited.
During sexual reproduction, the female germ cell will generally contribute more cytoplasm to the new organism. So, in cases of true cytoplasmic inheritance, differences in the inheritance of genes in reciprocal crosses are to be expected. However, the differences in the inheritance of genes in reciprocal crosses are not always indicative of true cytoplasmic inheritance. Predeterminations such as maternal inheritance and dauermodifications may also present differences in reciprocal crosses and therefore must be considered before cytoplasmic inheritance can be established as one of several modes of inheritance.
Maternal Inheritance Vs Cytoplasmic Inheritance
At first glance maternal inheritance will seem to be cytoplasmic inheritance. As mention before, the female germ cell will generally contribute more cytoplasm to a newly developed organism and occasionally, substances found in the female’s cytoplasm will influence the phenotype of the organism. As such it will appear as if the offspring is inheriting genes outside of the genes inherited in the nuclear inheritance model. A classic example of such inheritance is the coiling direction of Lymnaea peregra’s shell. The direction of coiling in offspring from dextral mothers is determined by maternal effect. The dextral gene produces a product that
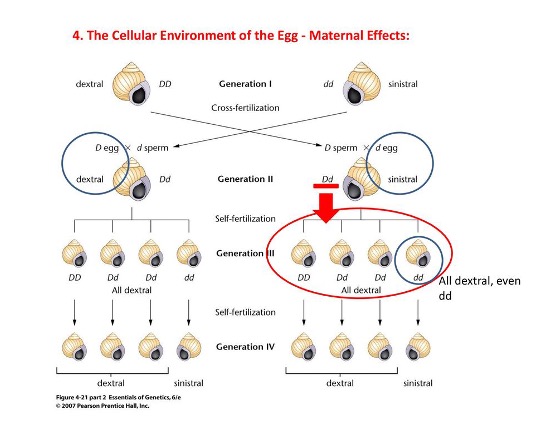
during oogenesis, influences the pattern of shell coiling in the embryo. Researchers have proved this by injecting into sinistral eggs, the cytoplasm from dextral mothers and saw the eggs from sinastral mothers exhibit a dextral coiling pattern.
It is easy to see why one may mistake this as cytoplasmic inheritance. After all, the phenotype of an offspring is be determined by factors that are not in the nuclear genome. However, it is very clear that this is not indeed cytoplasmic inheritance because it is not gene being inherited outside of the cytoplasm that is causing this phenotype, but properties of the cytoplasm inherited from the mother influencing the phenotype.
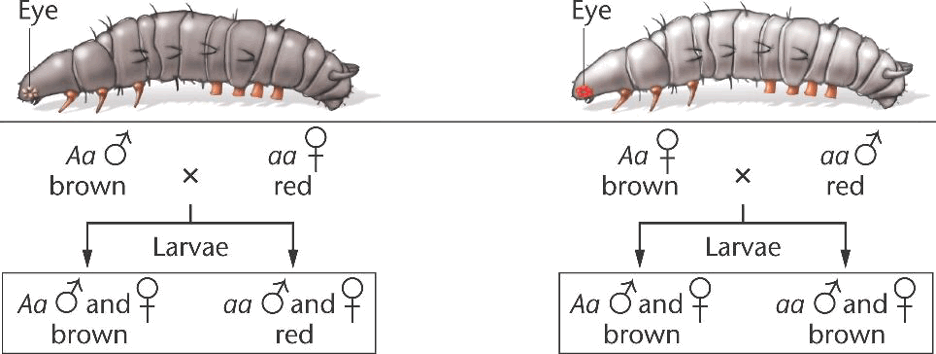
Another example, displaying the commonality of this phenomenon, occurs in Ephestia kuhniella. The recessive gene a that determines a reduction of pigment in the eyes, testes, and brain of this species. In crosses ♀ aa x ♂ Aa the distribution of each phenotype can be easily distinguished. But when the reciprocal cross occurs, all offspring initially display the Aa phenotype though later in life they are discovered to be of the aa phenotype. This occurs because in mothers with the Aa phenotype a substance known as kynurenin that is created by the Aa gene is stored in the egg and influences the phenotype of the offspring.
Dauermodification vs Cytoplasmic Inheritance
Dauermodifications are environmentally induced cytoplasmic changes transmitted maternally through successive generations. Dauermodifications have been documented by Jollos in 1913 and Sonneborn in 1943 in protozoa, in animals by Plough and Ives in 1935, and in plants by Hofmann in 1927 (Ernst 1948). In all these cases, phenotypes in the parental generation induced by environmental factors such as temperature and chemical exposure were inherited by the organism’s offspring. In all cases however the changes were temporary, and the conditions previous to the environmental changes were reverted to in successive generations.
Hofman in 1927 induced chloral hydrate leaf aberrations in Paseolus (Ernst 1948). This aberration was transferred through the ovum and not through the pollen and was noted to disappear after six generations. Sirks described a bud variation in Paseolus that appeared spontaneously and disappeared after eight generations of selfing.
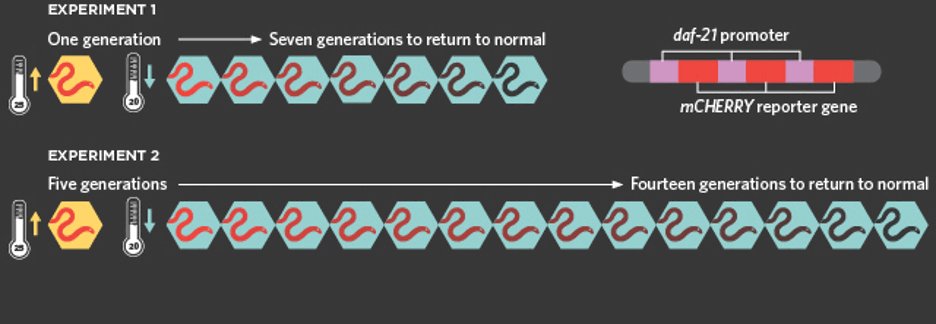
A direct example of dauermodification is in Nematodes. In 2017, researchers engineered Caenorhabditis elegans with numerous copies of transgene mCHERRY which is fostered by a promoter for the gene daf-21. Researchers kept the worms at 25 degrees Celsius and observed the worms turn fluoresce red. These worms then went on to have progeny that showed a similar phenotype, despite never having experience the higher temperature and lacking the nuclear genes for this specific phenotype. This continued for seven generations before the worms reverted to their original phenotypes. Furthermore, researchers also kept the worms at 25 degree Celsius for five generations and the inherited phenotype lasted longer (Offord 2017).
In all these cases was makes it possible to distinguish between dauermodifications and true cytoplasmic inheritance is the fact that the inherited phenotype disappeared over time. If an organism were experiencing an inheritance of a stable unit, the characteristics would persist over generations.
Cytoplasmic Inheritance in Eukaryotes
Eukaryotes exhibit variations in the way cytoplasmic inheritance is facilitated. In one kingdom, the animal kingdom, cytoplasmic inheritance is facilitated by the inheritance of mitochondria, the energy producers of the cell. In the plant kingdom, cytoplasmic inheritance is facilitated by the inheritance of mitochondria and plastids (not to be confused with plasmids; a circular structure that carries genetic information). Before, each mechanism is examined let us familiarize ourselves with the anatomy and physiology of plastids and mitochondria.
Plastids
Proplastids are an organelle located in the cells residing in the meristematic regions of a plant, it is here that plastids originate. As more proplastids are produced and maturation of the cell, the cell the transforms into plastids, a group of semiautonomous cells with specialized functions. It is important to note that plastids (along with mitochondria) are said to have originated from an endosymbiotic event between two cells millions of years ago, but that remains a discussion for another time.
Types of Plastids
Plastids come in many different forms each with a different specialized function. They are associated with many different activities in plants such as photosynthesis and metabolic reactions that provide the plant with energy. Here is a list of all the plastids discovered as of today (Aryal 2020).
- Chloroplasts-Chloroplasts are plastids that are in the mesophyll cells on plant leaves; this is the site of photosynthesis.
- Chromoplasts- Chromoplasts are brightly colored plastids that act as the sigte of pigment accumulation. They are typicalled found in the fleshy fruits, flowers as well as various other pigmented parts of the plant such as leaves.
- Gerontoplast – Gerontoplast are formed during senescence (the degradation of various organelles of a plant). This plastid has therefore been suggested to play an important role in controlling the degradation of the chloroplasts.
- Leucoplasts- Generally, leucoplasts are colorless plasmids that are commonly found in colorless leaves and rabidly growing tissues (tubers, stems, roots, etc). They serve as the site of starch formation and storage.
- Amyloplasts – Amyloplasts are a type of plasmid involved in long term storage of starch. Like the other plasmids, amyloplasts develop from proplasmids. DNA
- Eliaoplast (Lipoplasts)- elaioplasts are a type of leucoplast that contain oil. They serve to store oils and lipids which explain the small drops of fat found inside the plasmids. DNA
- Proteinoplasts – Proteinoplasts contain higher levels of protein as compared to the other plastids. The proteins either accumulate as amorphous or crystalline inclusions and bound by a membrane.
Despite the great variety of listed plastids, the structural make up of plastids are quite similar. All plastid features a double membrane made up of galactolipids and other lipids and proteins, an internal membrane which plays an important role in the transport of various materials into and out of the cytoplasm, and lastly the stroma, the internal space that is enclosed by the double membrane of the plastid. Here lies the gene that are inherited via the cytoplasmic inheritance model.
Cytoplasmic Inheritance via Plastid Inheritance
Plant offspring will inherit their plastids via an extranuclear mode of inheritance. The previous belief is that during cell division only the nuclear elements of the cell are passed on from parent to offspring. Plastids are generally inherited uniparentally, though there has been documented cases of biparental inheritances. The genes located in the plastids are replicated when these organelles duplicate, and the gene located in the plastids will be passed on from parent to offspring. It is through this mode of inheritance that plants will undergo cytoplasmic inheritance.
Cytoplasmic Inheritance of Pollen Sterility in Maize
Rhoades, in 1933 described the cytoplasmic inheritance of pollen sterility in Maize. After obtaining a male sterile line, he outcrossed to pollen from a non-sterile line of plants (Ernst 1948). The result, all offspring were sterile, showing no evidence of the segregation of male sterility. Furthermore, chromosomes from fertile maize where marked and subsisted for the corresponding chromosome in the male sterile lines and this still resulted in all offspring being male sterile. This test was conducted in all ten chromosomes of maize with the same result. Rhoades, then took advantage of the fact that male sterile plants sometimes produced a varying amount of pollen. Rhoades pollinated 49 of these pollen producing plants from the male sterile line with progeny from the male fertile line. The results? Forty-six of them gave varying percentages of male sterile offspring, while three gave normal offspring only. It was concluded that the phenotypically fertile strain must contain the necessary qualities for male sterility. The existence of the male sterile plants that produced viable pollen permitted the observation that factors responsibility for male sterility is not transmitted by the pollen. Their must be a cytoplasmic inheritance of the phenotype of male sterility.
Cytoplasmic Inheritance in Pathogenicity in Rusts
Cases of cytoplasmic inheritance has also been documented by Johnson, Newton and Brown in Puccina graminis, Mycosphaerella graminicola and Heterodera avenae, all types of funguses pathogenic to plants. The sexual stages of these rust are represented by the uninucleate pycniospores formed on the host. Both + and – sexes exist. Cells from single Puccinia are transferred to another infected leaf. If the Puccini are of reciprocal sexes they will copulate and reproduce to form binucleate aeciospores, the infective stage for the Puccina. Parentals with differing physiological differences towards the pathogenicity towards inbred strains of the host were crossed. In most cases, offspring exhibited the expected mendelian ratios, but in two cases reciprocal differences existed. In these few cases, the offspring exhibited the same pathogenic characteristics towards the host as their its parent. Since the pycniospores transferred are in such a small quantity, carrying minute amounts of cytoplasm, it is determined that pathogenicity is inherited via cytoplasm from maternal strain.
Cytoplasmic Inheritance Through Mitochondrial Inheritance
Discovered in 1940 by Carl Benda, mitochondria have been the subject of a lot a scientific scrutiny. As a result of all this scrutiny, we have learned a lot about mitochondria. For instance, the makeup and composition of mitochondria is widely known. Mitochondria is made up of a double membrane. It is here that entry and exit of the cell is regulated. Mitochondria also has an intermembrane space, located between the two membranes. The cristae which folds out from the intermembrane serves as to increase the surface area and is the site of ATP synthesis.
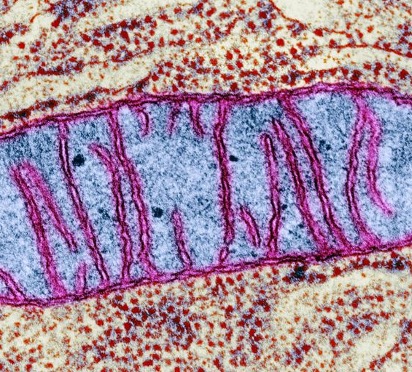
It is widely known that the function of mitochondria is to produce energy for the cell. Mitochondria breaks down food into simpler, charged molecules and combine them with oxygen to produce Adenosine Triphosphate (ATP), the energy currency of the cell. We also know that Mitochondria play a role in maintaining cellular metabolism by acting as mediators of signals that propagate various cellular processes. And recently it has been discovered that the mitochondria in cells may have originated from an endosymbiotic cell with an ancestral cell millions of years ago.
Significant, is the discovery that mitochondria carry its own DNA. What we find is that mitochondria contain circular DNA that codes for specific proteins and can replicate outside of nuclear control. In plants and humans, during sexual reproduction, the mitochondria are not broken down during cellular division. Instead, the mitochondria are inherited in the resulting offspring. It is during this extra nuclear inheritance of the mitochondria that plants and animals manifest cytoplasmic inheritance.
Cytoplasmic Inheritance in Humans
In the cases we have discussed already, it all concerned cytoplasmic inheritance in plants. However, as you could have guessed, cytoplasmic inheritance is also known to occur in animals. Here we will discuss the cytoplasmic inheritance in humans.
In humans, mitochondria are inherited via a uniparental mode of inheritance. All mitochondria are inherited strictly maternally It should be noted that during sexual reproduction spermatic mitochondria do not enter the oocyte cytoplasm and are not incorporated in embryo development (though in some extremely rare cases, offspring has also been shown to inherit their mitochondria maternally as well). In tandem, the mitochondrial DNA sequences are also inherited. It is this way that humans undergo cytoplasmic inheritance.
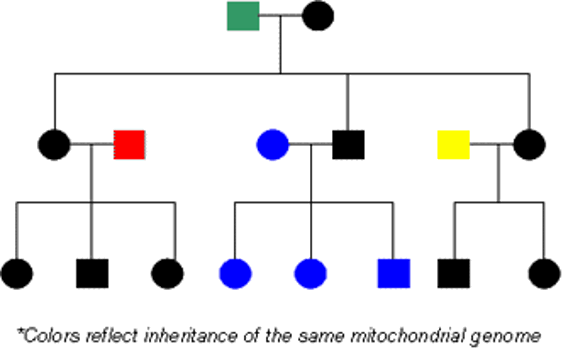
A key feature in the inheritance of mitochondrial DNA (mtDNA) is that mitochondrial DNA in offspring, is matroclinous like the mitochondrial DNA seen in previous generations. As a result, it is incredibly useful in the identification of members of relatives, even forensically.
However, like most mechanisms, cytoplasmic inheritance is subject to errors. These errors result in mitochondrial syndromes, a disease which occur due to the mutations of mitochondrial DNA (mtDNA). These mutations can be widespread, or isolated to specific tissue. As of today, there is no cure to mitochondrial syndrome, but research remains on going.
Cytoplasmic Inheritance in Habroibracon
Worthy of note is the early research that established a model for cytoplasmic inheritance in animals. In 1927, A. Kuhn selected two strains of Habrobracon from wild material. In one strain, the phenotype was darker than the other (Ernst 1948). The first generation of females’ phenotype was intermediate and those from lighter mothers were far lighter that females in reciprocal crosses. This difference was noted to be maintained in both the daughters produced in backcrosses to males of the light strain and haploid sons produced by the hybrid females. The same differences were noted in the sons produced by the females from the backcross and the ones from the reciprocal. It was concluded by Kuhn that the cytoplasm of the light strain would cause a lighter phenotype than other offspring with the same genotype.
Emerging Discoveries in Cytoplasmic Inheritance
Cytoplasmic Inheritance may appear to follow a random pattern of inheritance, but as we study such mechanisms, new patterns of inheritance emerge that reveal a diversity of complex processes. We have discovered that mtDNA has a close relationship with genes located in the nucleus. So much so, that the very integrity of the mtDNA may depend on the actions of the nuclear genes. Being that we are still discovering new details about cytoplasmic inheritance, cytoplasmic inheritance is still categorized as a mechanism that we are only just now beginning to by understood.
Conclusion
Along with nuclear inheritance, cytoplasmic inheritance interacts to influence heredity. The complexity of the inner workings of these parts are relevant even today to significant effects because of its many wide-reaching implications. Life as we know it, natural selection as we know it, are all guided by these principles and the principles of nuclear inheritance.
Citations
Aryal S. 2019. Plastids- Definition, Structure, Types, Functions and Diagram. Online Microbiology Notes, February 15, 2020. https://microbenotes.com/plastids-types-structure-and-functions/.
Caspari, E. 1948. “Advances in Genetics”. Academic Press Inc., Publishers New York, N.Y.,
Vol. 2 pages 1-61 February 10, 2020. https://www.sciencedirect.com/science/article/abs/pii/S0065266008604654?via=ihub.
Chial H. 2008. MtDNA and mitochondrial diseases. Nature News 217, Nature Publishing Group,
Febuary 21, 2020. https://www.nature.com/scitable/topicpage/mtdna-and-mitochondrial-diseases-903/#.
Freeman G, and Lundelius J. 1971 The Developmental Genetics of Dextrality and
Sinistrality in the Gastropod Lymnaea Peregra. Development Genes and Evolution. Wilhelm Roux Archives 191, 69-83, February 15, 2019. https://link.springer.com/article/10.1007/BF00848443.
Hagemann, R. 2000. Erwin Baur or Carl Correns: Who Really Created the Theory of Plastid
Inheritance? OUP Academic. Oxford University Press, Pages 435-440. February 15, 2020. https://academic.oup.com/jhered/article/91/6/435/2187039.
Hagemann, R. 2010. The Foundation of Extranuclear Inheritance: Plastid and Mitochondrial
Genetics. Molecular Genetics and Genomics 283 (3) (03): 199-209. February 16, 2020 http://ezproxy.purchase.edu:2048/login?url=https://search.proquest.com/docview/194164737?accountid=14171.
Herschy, Barry. Natures Electrochemical Flow Reactors?: Alkaline Hydrothermal Vents and the
Origins of Life. The Biochemist 36, no. 6 (January 2014): 4–8. https://doi.org/10.1042/bio03606004.
Inaba T, Fumiko Y, Yasuko I, Tomohiro K, and Katsuhiro N. 2011. Retrograde Signaling
Pathway from Plastid to Nucleus. International Review of Cell and Molecular Biology Vol 290,2011, Pages 167-204. Academic Press, Febuary 20, 2020. https://www.sciencedirect.com/science/article/pii/B9780123860378000028.
Mandal, Ananya. Gene History. News, April 22, 2019. https://www.news-medical.net/life-sciences/Gene-History.aspx.
Miko, I. 2008. Non-Nuclear Genes and Their Inheritance. Nature News. Nature Publishing
Group 1(1) 135. February 10, 2020. https://www.nature.com/scitable/topicpage/non-nuclear-genes-and-their-inheritance-589/#.
Offord C. 2017 Epigenetic Inheritance in Nematodes. The Scientist
Magazine®. July 16, 2017. https://www.the-scientist.com/the-literature/epigenetic-inheritance-in-nematodes-31228
Olby, R, 2020. Gregor Mendel. Encyclopædia Britannica. Encyclopædia Britannica, inc., January 2, 2020. https://www.britannica.com/biography/Gregor-Mendel.
Orr, A. Darwin and Darwinism: The (Alleged) Social Implications of The Origin of Species. Genetics. Genetics Vol. 183 no 3, November 1, 2009. https://www.genetics.org/content/183/3/767.
Salam, Kamoru A. Towards sustainable development of microalgal biosorption for treating effluents containg heavy metals. Biofuel Research Journal 6, no. 2 (2019): 948–61. https://doi.org/10.18331/brj2019.6.2.2.
Sato N, Kimihiro T, Kazunori M, and Yukihiro K. 2004. Organization,
Developmental Dynamics, and Evolution of Plastid Nucleoids. International Review of Cytology Vol 232,2003, Pages 271-262. Academic Press, Febuary 16, 2020. https://www.sciencedirect.com/science/article/abs/pii/S0074769603320066.
Takahashi, H, and Kan T. 2014 Intergenomic Transcriptional Interplays between Plastid as a
Cyanobacterial Symbiont and Nucleus. Progress in Biotechnology. Elsevier Science B.V. Vol 22,2002, Pages 105-120, Febuary 16, 2020. https://www.sciencedirect.com/science/article/abs/pii/S0921042302800478.
Yurtoğlu, Nadir. “Http://www.Historystudies.net/dergi//birinci-Dunya-Savasinda-Bir-Asayis-
Sorunu-Sebinkarahisar-Ermeni-isyani20181092a4a8f.Pdf.” History Studies International Journal of History 10, no. 7 (2018): 241–64. https://doi.org/10.9737/hist.2018.658.



{kind=link}
More Stories
Water Bears, Earths Mightiest Creatures!